Er cellen en maskin, eller mer som et sinn?
Av Robert Shedinger; 7. sept. 2023. Oversatt herfra
Bilde 1. Kinesin som frakter sin last
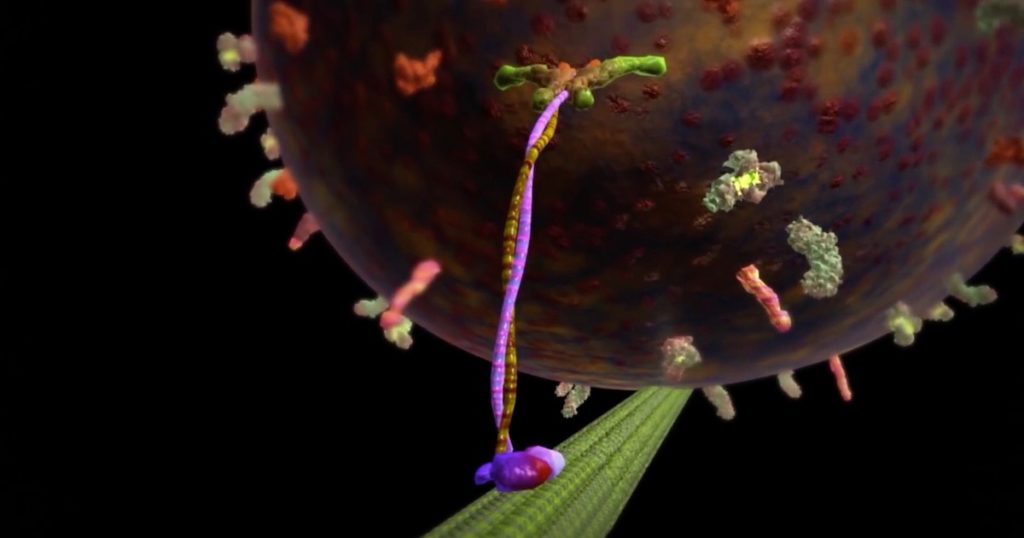 I 2019 dukket det opp en artikkel i Journal of Theoretical Biology og spurte "Er cellen virkelig en maskin?" Forfatteren, Daniel Nicholson fra Konrad Lorenz Institute for Evolution and Cognition Research, argumenterer mot det ofte holdte synet på cellen som maskinlignende.1 Hvis ikke en maskin, hva da? I det minste som vi er vant til å tenke i vår KI -tidsalder, er alternativet til en maskin et sinn. åpner Nicholson da muligheten for at cellen er mer som et intelligent sinn? Han ignorerer den ideen totalt, som er ironisk gitt at han jobber på et institutt dedikert til kognitivitetsforskning. Men la oss ta en titt.
I 2019 dukket det opp en artikkel i Journal of Theoretical Biology og spurte "Er cellen virkelig en maskin?" Forfatteren, Daniel Nicholson fra Konrad Lorenz Institute for Evolution and Cognition Research, argumenterer mot det ofte holdte synet på cellen som maskinlignende.1 Hvis ikke en maskin, hva da? I det minste som vi er vant til å tenke i vår KI -tidsalder, er alternativet til en maskin et sinn. åpner Nicholson da muligheten for at cellen er mer som et intelligent sinn? Han ignorerer den ideen totalt, som er ironisk gitt at han jobber på et institutt dedikert til kognitivitetsforskning. Men la oss ta en titt.
Jeg er ikke i stand til å felle dom om det tekniske i molekylærbiologi, presentert av Nicholson; Jeg overlater den vurderingen til andre. Men hvis artikkelen hans i det hele tatt er vitenskapelig nøyaktig, blir språket han bruker for å beskrive cellulære prosesser, gjennomvevd med implikasjoner av cellulær kognisjon, utover alt som er tenkt av Barbara McClintock -lenke. I to innlegg vil jeg analysere Nicholsons språkbruk.
Helt siden Monod
Nicholson sporer det moderne maskinlignende synet på cellen tilbake til arbeidet til Jacques Monod og hans innflytelsesrike Chance and Necessity. Helt tiden siden, ifølge Nicholson, har molekylærbiologi tatt i bruk et tolkende rammeverk der cellen forstås som:
"Et intrikat maskineri hvis organisasjon gjenspeiler en eksisterende design, hvis struktur er forståelig i reduksjonistiske termer, og hvis operasjon styres av deterministiske lover, noe som gjør atferden forutsigbar og kontrollerbar-i det minste i prinsippet (108)."
Nicholson kaller denne "MaskinForestillingrm av Cellen" eller MFC. Under dens innflytelse har metaforiske termer som låser, nøkler, porter, pumper og kretsløp kommet til å gjennomsyre språket i molekylærbiologi.
Nicholson synes dette er underlig, for etter hans syn har moderne forskning skapt et syn på cellen helt i strid med denne maskinlignende unnfangelsen. Hans analyse dreier seg om fire spesifikke aspekter av cellen: cellulær arkitektur, proteinkomplekser, intracellulær transport og cellulær atferd. I begge tilfeller introduserer Nicholson ideer om selvorganisering og stokastisitet for å undergrave reduksjonistiske og deterministiske forestillinger om cellulær atferd. Men i prosessen ser han ut til å bringe cellulær intelligens inn i bildet. Jeg vil vurdere hvert av de fire eksemplene i detalj.
Cellulær arkitektur
 I følge Nicholson har celler tradisjonelt blitt forstått som oppstått gjennom prosessen med selvmontering, definert som:
I følge Nicholson har celler tradisjonelt blitt forstått som oppstått gjennom prosessen med selvmontering, definert som:
"Den fysiske assosiasjonen av molekyler til en statisk likevektsstruktur i fravær av en ekstern energikilde. Det er drevet av lokale stereospesifikke interaksjoner mellom de aggregerte 'byggesteinene', som forblir uendret gjennom hele prosessen (110)."
Derimot foretrekker Nicholson den mer dynamiske ideen om selvorganisering, hvorved en samling av molekyler kan opprettholde seg selv i en langt fra likevektstilstand ved å "stadig bruke energi og utveksle materie med omgivelsene" (110). I dette mer dynamiske synet er cellen bedre karakterisert som en "metastabil fluks dynamisk reagerer på endringer i miljøet enn som en statisk makromolekylstruktur" (111).
Bilde 2. Om biologiens seier over kjemi (ironi innebygd)
Maskinmetaforen autoriserer et bilde av cellen som statisk strukturert av et genetisk program, omtrent som maskiner er statisk strukturert av en blåkopi. Men selvorganisering endrer grunnleggende denne forestillingen:
Den selvorganiserende naturen til den cellulære arkitekturen har vidtrekkende teoretiske konsekvenser. Mest grunnleggende fører det til et syn på cellen som er helt i strid med MFC. For det første fordriver den forestillingen om at ‘informasjonen’ som spesifiserer den romlige organisasjonen av cellen på en eller annen måte er kodet i genomet. Strengt tatt er det ingen genetisk blåkopi for celle- arkitekturen. Selvorganisering genererer orden i mangel av en ekstern mal eller global plan (112).
Så ifølge Nicholson er cellulær arkitektur i en konstant flukstilstand da cellulære komponenter kontinuerlig samhandler på måter å få til de arkitektoniske strukturer som er nødvendige i hvert øyeblikk for å svare på øyeblikkets behov, hvoretter disse strukturene forsvinner for å erstattes av nye. Men hvordan kan dette nivået av dynamikk ved forklaring uten appell til et visst nivå av cellulær erkjennelse? Hvis cellulær arkitektur ikke er statisk og spesifisert av en forhåndsbestemt genetisk plan, men snarere er i en dynamisk flukstilstand som alltid reagerer på riktig måte på cellens øyeblikkelige behov, ser det ut til at intelligens er sentral i cellulær atferd ("atferd" i seg selv er en kognitiv idé).
Interessant nok spør Nicholson hvorfor celler vil favorisere selvorganisering fremfor selvmontering, noe som antyder at celler har et bevisst valg i saken! "Ville det ikke," spør han, "gi mer mening for en celle for å bygge statiske, likevektsstrukturer som ikke krever en konstant energiutgifter for å opprettholde dem?" (112). Mens han erkjenner at selvmontering ville være mer økonomisk og effektiv, konkluderer han med at selvorganisering "lar celler svare raskt og tilpasse seg til eksterne forstyrrelser og andre kritiske hendelser som ellers ville sette deres systemiske integritet i fare" (112). Men hvis en celle kan velge selvorganisering for sin større adaptive fleksibilitet, må cellen være en slags intelligent agent. å favorisere en ting fremfor en annen er en sinnsaktivitet, uansett.
Bilde 3. Et høne-egg problem mellom proteiner og enzymer/membraner
Proteinkomplekser
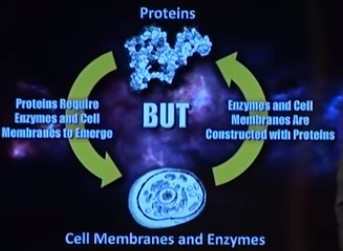 MFC har ifølge Nicholson ført til ideen om at proteinkomplekser representerer faste enheter hvis struktur bestemmes av aminosyresekvenser spesifisert i DNA-koden, og som utfører maskinlignende funksjoner i cellen bestemt av deres unike tredimensjonale form . Men nyere forskning antyder at proteinkomplekser er langt mer flytende og dynamiske.
MFC har ifølge Nicholson ført til ideen om at proteinkomplekser representerer faste enheter hvis struktur bestemmes av aminosyresekvenser spesifisert i DNA-koden, og som utfører maskinlignende funksjoner i cellen bestemt av deres unike tredimensjonale form . Men nyere forskning antyder at proteinkomplekser er langt mer flytende og dynamiske.
Nicholson peker på det kuleproteinlymfotaktinet, som ikke har noen fast konformasjon, men heller gjennomgår store strukturelle endringer når det flimrer frem og tilbake mellom to forskjellige konformasjoner. Så er det den nylige oppdagelsen av iboende forstyrrede proteiner (IDP) som ikke har noen bestilte konformasjoner, men i stedet "streifer rundt i cellen som utfoldede polypeptid-kjeder" (114). IDP -er gir en distinkt funksjonell fordel fordi de er i stand til å "samhandle med et bredt spekter av bindingspartnere (inkludert andre proteiner, membraner, nukleinsyrer og forskjellige mindre molekyler) ved å ta i bruk forskjellige konfigurasjoner" (114). I stedet for å ha en fast funksjon basert på en fast tredimensjonal form, bestemmes funksjonen til disse proteinene av miljøet og interaksjonene de har med molekylene rundt seg. "Funksjonell promiskuitet," som Nicholson betegner det, ser ut til å være regelen snarere enn unntaket for proteiner.
Dessverre har Nicholson ingenting å si om hvordan proteiner utfører dette trikset. Hvis det ikke er noen kjemisk nødvendighet som driver de forskjellige funksjonene som er tatt opp av disse fluidproteinene, hvordan vet proteiner hvilken konformasjon og funksjon de skal ta i bruk til enhver tid? Igjen ser dette ut til å innebære muligheten for et visst nivå av kognitiv evne på molekylernivå. Kanskje var Sewall Wright ikke så langt unna da han spekulerte i hvordan det føles å være et hydrogenion! (2) Har proteiner et indre liv?
I dette avsnittet tar Nicholson også problem med bruk av ledningsdiagrammer for å skildre kompleksiteten i cellulære kretsløp. Etter hans syn gjør proteinerens dynamiske natur alle faste ledningsdiagrammer foreldet. Kanskje en bedre metafor er nevralt i stedet for elektronisk kretsløp. Deretter vil vi vurdere hva Nicholson har å si om intracellulær transport og cellulær oppførsel.
 For referanser se slutten av originalartikkelen -lenke.
For referanser se slutten av originalartikkelen -lenke.
Bilde 4. Robert F. Shedinger
ROBERT SHEDINGER (bildet)
Robert F. Shedinger er professor i religion ved Luther College i Decorah, Iowa. Han har en Ph.D. i religionsvitenskap fra Temple University og er nylig
forfatter av The Mystery of Evolutionary Mechanisms: Darwinian Biology's Grand Narrative of Triumph and the Subversion of Religion.
Han underviser jevnlig på kurs om forholdet mellom vitenskap og religion.
Oversettelse og bilder ved Asbjørn E. Lund